International Journal of Pharmaceutical and Phytopharmacological Research
ISSN (Print): 2250-1029
ISSN (Online): 2249-6084

Antimicrobial resistance (AMR) occurs when microorganisms fail to respond to the therapeutic onslaught of antibiotics. Extended-spectrum beta-lactamase (ESBL) and AmpC enzymes are important AMR mechanisms that erode the efficacy of important antibiotics. Here, we report on the detection and susceptibility of ESBL- and AmpC-producing bacteria from livestock and poultry environments. Bacteriological and molecular biology tools were used for the isolation and characterization of bacteria. Combined disk diffusion methods and PCR were used to screen and confirm ESBL and AmpC production. ESBL was phenotypically detected in E. coli, Klebsiella species, and P. aeruginosa for samples from poultry at the rate of 4%, 1%, and 2% while samples from livestock milieus had ESBL-positive bacteria at the rate of 5%, 2%, 4% for E. coli, Klebsiella species and P. aeruginosa respectively. AmpC was phenotypically detected in E. coli (3%), Klebsiella species (2%), and P. aeruginosa isolates (1%) for samples from poultry milieus. For samples from livestock milieus, AmpC was phenotypically detected in E. coli (7%), Klebsiella species (3%), and P. aeruginosa (6%). The ESBL- and AmpC-positive bacteria showed significant levels of reduced susceptibility to the carbapenems and cephalosporins. PCR detected CTX-M-15 genes (20%) and FOX-1 genes (25%) which mediated ESBL and AmpC resistance in bacteria. These findings have led to the identification of key functional genes that cause bacterial resistance in southeast Nigeria, and focus attention on the importance of surveillance and monitoring to mitigate the transmission of AMR in the environment, as antibiotic therapy could be affected.
INTRODUCTION
Antimicrobial resistance (AMR) is a significant health menace of this century and impacts developing healthcare systems around the world. It involves the evolution, mobilization, and transfer of AMR bacteria and antibiotic resistance genes (ARGs) between humans, animals, and the environment; thereby reducing our ability to effectively treat common microbial (bacterial) infections. Members of the Enterobacteriaceae family and non-enteric bacteria are currently the most clinically important group of bacteria responsible for AMR-related deaths worldwide, in the food chain and hospital settings in high- and low- and middle-income countries contexts. AMR can be mobilized from the community or hospital environment through mobile genetic elements (MGEs) including plasmids and transferred through genetic transfer mechanisms (e.g., conjugation) to other bacteria in the same milieu. The presence of this AMR bacterium can favor the acquisition and maintenance of these MGEs that stimulate the evolution and spread of ARGs. Recent literature has estimated that by 2050, 10 million deaths per year will be attributable to AMR worldwide [1]. The widespread and irrational usage of antibiotics, particularly in livestock and poultry endeavors contributes to the evolution and transmission of AMR/ARGs in the community and hospital environment. Extended-spectrum beta-lactamases (ESBLs) and AmpC enzymes are important resistance enzymes that give pathogenic bacteria the advantage to ward off the antimicrobial activity of some antibacterial agents. The increasing prevalence of these resistant bacteria and ARGS is of public health importance since the presence of AMR bacteria limits the choice of effective antibacterial agent for therapy [2-6]. ESBLs and AmpC enzymes confer bacterial resistance to the second and third- generation cephalosporins – which are important antibacterial agents used clinically for treating bacterial-related infections [5-7]. Both ESBLs and AmpC enzymes give bacterial pathogens such as P. aeruginosa, E. coli, and Klebsiella species the extra advantage of resisting the killing or inhibitory potentials of some available antibacterial agents, consequently making it challenging to achieve optimal antimicrobial therapy and prognosis in infected patients. Resistant P. aeruginosa, E. coli, and Klebsiella species are among the priority-resistant microorganisms implicated in bacterial-related infections occurring in the community or hospital environment globally [7-9]. The reduced susceptibility of pathogenic bacteria to antimicrobial agents is a global phenomenon spanning from one country to another, and signifies a chief and severe public health menace in hospitals across the world, thus making it difficult for physicians to select and choose the best antibiotic for managing some infections caused by these pathogens. This is because some of these resistant pathogenic bacteria are already showing multidrug resistance to a wide variety of available antibiotics used in clinical medicine [2, 10-12]. Since antibiotics are not completely degraded upon usage, and resistant bacteria can show up in food and other sources including the food chain, and could be a major route for the transmission of AMR/ARGs in any environment, it is important to monitor and report the increasing prevalence of these multidrug-resistant bacteria producing ESBL and AmpC in the non-hospital environment as a way of assuaging the public health risk that they may pose in human populations. An understanding of the threats and mobilization of AMR/ARGs from the non-hospital environment and their impact on human health is essential for improving current AMR protocol based on gained experience, as well as mitigating the evolution, persistence, and transmission of AMR/ARGs in these milieus and human population. It is against this backdrop that this current research examined samples from outside the hospital environment for the spread of these AMR/ARGs, particularly ESBL and AmpC genes in P. aeruginosa, E. coli, and Klebsiella species from livestock and poultry milieus in southeast Nigeria.
MATERIALS AND METHODS
Sampling, culture, isolation, and identification of bacteria
Ethical approval for this research was sought from the Local Ethics Committee of Enugu State University of Science and Technology (ESUT), Agbani, Nigeria prior to the start of the study. The protocols in this study were carried out in line with relevant national rules concerning research involving animal samples. A total of 150 and 300 non-duplicate samples from livestock and poultry environments were collected over a nine (9) months period [January 2021 – September 2021] from various animal farms in southeast Nigeria for this present study. All samples included in this study were processed using standard microbiology techniques for the isolation of Gram-negative bacteria that included E. coli, Klebsiella species, and P. aeruginosa on their respective selective culture media [13-15].
Screening of bacterial isolates for ESBL and AmpC enzymes
ESBL and AmpC enzymes were phenotypically detected in the test E. coli, Klebsiella and Pseudomonas aeruginosa isolates using the combined double disk diffusion technique as per our previous method [2, 3, 13]. Based on this protocol, all test isolates were swabbed on Mueller Hinton (MH) agar plates and subjected to the requisite antibiotic disks, particularly the third-generation cephalosporin (for ESBL detection) and second-generation cephalosporins (for AmpC enzyme detection). A ≥5mm increase in the inhibition zone diameter for any of the third-generation cephalosporin (ceftazidime and cefotaxime) tested in combination with AMC (20/10µg) compared to its inhibition zone diameter when tested alone was inferred as ESBL positive phenotypically. For AmpC enzyme production, AmpC phenotypes were phenotypically inferred when the test bacterial isolates showed obvious blunting or flattening of the zones of inhibition between the ceftazidime (CAZ), imipenem or cefotaxime (CTX) disks adjacent to the cefoxitin disk.
PCR typing of the resistance genes
Plasmid DNA was extracted from the test bacterial isolates using the Zymo Plasmid miniprep kit (Epigenetics Company, USA). Table 1 shows the primers that were used for the PCR amplification of ESBL and AmpC genes. The purified DNA fragment was quantified using a NanoDrop (Thermo Scientific, USA). PCR experiment was conducted according to the procedures described previously [16, 17]. This was carried out using a PCR master mix (50 µl) comprising 5x GoTaq (10µl), 25 mM MgCl2 (3µl), dNTPs (10 mM) 1µl, forward primers (1µl), reverse primers (1µl), 10 pmol 1 µl, DNA Taq (1000 U) 25µl, Ultrapure Water 8µl. The PCR conditions used are an amplification cycle at 95 °C for 5 minutes; 35 cycles at 96 °C for 30 seconds, 58 °C for 90 seconds, and 72 °C for 60 seconds; and a final extension step at 72 °C for 10 minutes.
Table 1. Oligonucleotide primers used for PCR experiment
|
Primer Name |
Primer Sequence direction |
Amplicon Size (bp) |
|
FOX-1 F |
AACATGGGGTATCAGGGAGATG |
190 |
|
FOX-1 R |
CAAAGCGCGTAACCGGATTGG |
190 |
|
blaTEM F |
ATGAGTATTCAACATTTCCG |
445 |
|
blaTEMR |
CCAATGCTTAATCAGTGAGC |
445 |
|
blaCTX-M-15 F |
CCCATGGTTAAAAAATCACTG |
850 |
|
blaCTX-M-15 R |
CCGTTTCCGCTATTACAAAC |
850 |
Key: F = Forward, R = Reverse
Antimicrobial susceptibility testing
Susceptibility studies were performed on all ESBL- and AmpC- producing bacterial isolates screened and isolated in this study. This was done using our previous method and in tandem with the modified Kirby-Bauer disk diffusion technique on Mueller Hinton agar (Oxoid, UK) plates according to the criteria of the Clinical Laboratory Standards Institute (CLSI) and according to a previous methodology [2, 18, 19]. Single antibiotic disks comprising: doxycycline (30μg), colistin sulfate (10μg), aztreonam (30μg), ceftazidime (30μg), ceftriaxone (30μg), cefotaxime (30μg), cefoxitin (30μg), sulphamethoxazole-trimethoprim (5μg), imipenem (10μg) and mupirocin (300μg) [Oxoid, UK] were used for susceptibility studies. All test bacterial isolates of ESBL- and AmpC-positive Gram-negative bacteria (adjusted to 0.5 McFarland turbidity standards) were aseptically swabbed on MH agar plate(s) and incubated at 37oC overnight. The results of the antimicrobial susceptibility studies were read and inferred as resistant and susceptible according to the criteria of CLSI after comparing the data obtained to the CLSI standard antibiotic breakpoints [18].
RESULTS AND DISCUSSION
We investigated the prevalence and antimicrobial susceptibility profiles of Gram-negative bacteria including E. coli, Klebsiella species, and P. aeruginosa isolates recovered from livestock and poultry milieus, as well as their ability to produce some antibiotic resistance factors, particularly, ESBL and AmpC enzymes, over a nine (9) months period in the southeast part of Nigeria. Both enteric and non-enteric bacteria were recovered from the livestock and poultry samples (Table 2).
Table 2. Isolation of enteric (E. coli, Klebsiella species) and non-enteric bacteria (P. aeruginosa)
|
Source |
Klebsiella species n(%) |
P. aeruginosa n(%) |
E. coli n(%) |
|
Poultry |
8 (3) |
5 (2) |
42 (14) |
|
Livestock |
13 (9) |
6 (4) |
35 (23) |
The most prevalent bacteria isolated from the test samples were E. coli which occurred at a rate of 23% and 14% for livestock and poultry samples respectively. This was followed by Klebsiella species, and then P. aeruginosa (Table 2). Table 3 shows the prevalence of ESBL- and AmpC-producing bacteria detected by phenotypic techniques from the livestock and poultry samples investigated in this study. ESBL was phenotypically detected in isolates of E. coli (4%), Klebsiella species (1%), and P. aeruginosa (2%) for poultry samples while AmpC enzymes were phenotypically detected in E. coli (3%), Klebsiella species (2%) and P. aeruginosa (1%) isolates (Table 3). Furthermore, ESBL was also detected in E. coli (5%), Klebsiella species (2%), and P. aeruginosa (4%) isolates emanating from poultry samples. For livestock samples, AmpC enzymes were phenotypically detected in E. coli (7%), Klebsiella species (3%) and P. aeruginosa isolates.
Table 3. Prevalence of ESBL and AmpC-positive bacteria by phenotypic finding
|
Bacteria |
ESBL n(%) |
AmpC positive n(%) |
ESBL n(%) |
AmpC positive n(%) |
|
|
Poultry (n=300) |
Livestock (n=150) |
||
|
E. coli |
12 (4) |
10(3) |
8 (5) |
11 (7) |
|
Klebsiella species |
2(1) |
5(2) |
3 (2) |
4 (3) |
|
P. aeruginosa |
5(2) |
3(1) |
6 (4) |
9 (6) |
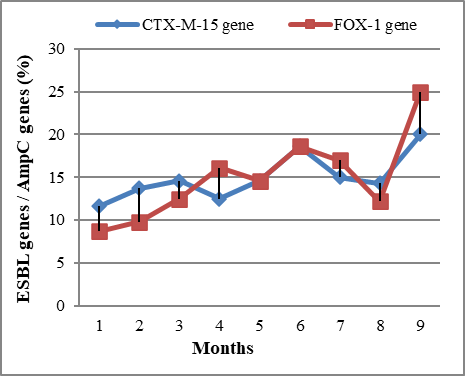
Surprisingly, the frequency of AMR mediated by ESBL and AmpC genes in the test bacterial isolates of E. coli, Klebsiella species, and P. aeruginosa, increased over the 9-month period of the study (Figure 1).
|
|
|
Figure 1. Preliminary results showing the rate of resistance mechanisms in the last year [Jan. 2021 – Sept. 2021]. Here we showed, over 9 months in southeast Nigeria, there was a rapid rise in AMR patterns of bacteria caused. |
Data shows a rapid rise in AMR mediated by ESBL and AmpC genes over 9 months in southeast Nigeria. It was observed in this study that the most prevalent ESBL gene that mediated AMR in the test bacterial isolates of E. coli, Klebsiella species, and P. aeruginosa is the CTX-M-15 gene – which is known to mediate bacterial resistance to cefotaxime, important third-generation cephalosporins used for the management of many bacterial infections in humans. Furthermore, the FOX-1 gene was found to be the most prevalent AmpC gene that was detected by PCR in the test bacterial isolates of E. coli, Klebsiella species, and P. aeruginosa (Figure 1). This gene (FOX-1) is known to mediate bacterial resistance to the second generation of cephalosporins – which are clinically relevant for the treatment of many bacterial-related infections in the human population. Figure 2 shows the result of antimicrobial susceptibility patterns of the ESBL- and AmpC-producing E. coli, Klebsiella species, and P. aeruginosa isolate to some selected clinically important antibiotics.
|
|
|
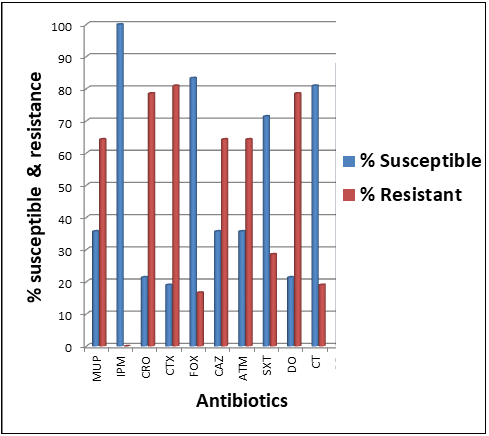
Figure 2. Chart showing antibiogram of the ESBL- and AmpC-producing bacterial isolates. Key: SXT = sulphamethoxazole-trimethoprim, DO = doxycycline, CT = colistin sulphate, ATM = aztreonam, CAZ = ceftazidime, CRO = ceftriaxone, CTX = cefotaxime, FOX = cefoxitin, IPM = imipenem, MUP = mupirocin |
The ESBL- and AmpC-producing bacterial isolates of E. coli, Klebsiella species, and P. aeruginosa from the livestock and poultry environments showed varying rates of susceptibility and resistance to the tested antibiotics. Interestingly, a high rate of resistance by the bacterial isolates was shown against important clinically relevant third-generation cephalosporins, particularly cefotaxime (81%) and ceftriaxone (78%). This was followed by mupirocin (64%), doxycycline (78%), ceftazidime (63%), and aztreonam (62%) to which the test bacterial isolates also showed reduced susceptibility (Figure 2). In all, imipenem (99%), cefoxitin (82%), and colistin sulfate (80%) were the most effective antibacterial agents against the ESBL- and AmpC-producing bacterial isolates recovered in this study.
Antimicrobial resistance (AMR) is a public health menace that compromises infectious disease control across the globe, and developing health systems are more fragile to the effects of increasing AMR. It knows not the border of any nation and can be transmitted from one place to another if sustainable mitigation strategies are not effectively implemented to alleviate and curtail their menace while preserving the clinical efficacy of available antimicrobial agents. Since extensive use of antibiotics in agriculture (e.g. animal husbandry) impacts public health and the environment in terms of the evolution and spread of AMR bacteria – which cause drug-resistant infections in the human population, we set out to evaluate how this phenomenon occurs in some farms in southeast Nigeria. Given this convincing situation, this study investigated how ESBL- and AmpC-producing E. coli, Klebsiella species, and P aeruginosa are transmitted in the livestock and poultry environments in southeast Nigeria by reporting on the prevalence of these important antibiotic-degrading enzymes in some selected non-hospital environments. The prevalence of the isolated E. coli, Klebsiella species, and P. aeruginosa isolate recovered from the poultry and livestock samples investigated in this study ranged between 3-23% - indicating an increase from our previously reported study on the frequency of enteric and non-enteric bacteria of clinical importance in the non-hospital environments [2, 4]. The isolated bacteria are important priority antibiotic-resistant bacteria recognized by the World Health Organization (WHO), and responsible for a handful of infections that warrants hospital visits in most parts of the world especially in developing healthcare systems where primary healthcare is still in a pitiable state. ESBL- and AmpC-producing E. coli, Klebsiella species, and P. aeruginosa were detected phenotypically at the rates of 4%, 1%, 2%, and 3%, 2%, 1% for samples that emanated from poultry milieus respectively. This evidence would suggest that ESBL- and AmpC-producing bacteria may be spreading in the animal and human populace which could result from the unmonitored and irrational use of antibiotics in poultry farms. For samples resulting from livestock environments, ESBL and AmpC-producing E. coli, Klebsiella species, and P. aeruginosa was phenotypically detected at the rates of 5%, 2%, 4%, and 7%, 3%, and 6% respectively. The prevalence of ESBL- and AmpC-producing E. coli, Klebsiella species and P. aeruginosa isolates by phenotypic detection reported in this study, from poultry and livestock environments is lower than our previous report in which ESBL- and AmpC-producing E. coli, Klebsiella species, and P. aeruginosa isolates were in the range of 10-30%. This report is lower than a related study in which ESBL- and AmpC-producing bacteria were highly reported in enteric and non-enteric bacteria from clinical environments in Iran [17] and India [16]. In New Zealand, a similar prevalence of ESBL-producing bacteria was reported in Enterobacteriaceae including E. coli and Klebsiella species [20]. The predominant type of ESBL- and AmpC genes detected by PCR, and which are occurring in the test isolates of E. coli, Klebsiella species, and P. aeruginosa were CTX-M-15 and FOX-1 genes respectively. CTX-M-15 is a globally distributed ESBL gene that mediates bacterial hydrolysis of third-generation cephalosporins while FOX-1 is an AmpC gene that mediates bacterial hydrolysis of the cephalosporins such as cephamycins [6, 12], thereby making antibiotics in this class to be therapeutically ineffective upon clinical application [21-25]. The ESBL- and AmpC- the activity of the test bacterial isolates of E. coli, Klebsiella species, and P. aeruginosa from both livestock and poultry environments in this study is in concordant with previous studies from the Czech Republic and Italy which indicate the carriage and spread of CTX-M ESBL genes and AmpC genes in the community [26, 27]. This may portend important clinical implications in terms of acting as a risk factor for the acquisition and transmission of ESBL- and AmpC-gene-encoding bacterial strains from these sources (animals) to humans. On the other hand, the prevalence of ESBL and AmpC-gene encoding strains from another study is between 10-40% [28-30]. Also in another related study, the frequency of ESBL bacteria in Africa ranges between 6.7-36.1 %, a report that slightly differs from ours [29]. The detection of CTX-M-15 ESBL gene and FOX-1 AmpC gene in approximately 20% and 25% of the E. coli, Klebsiella species and P. aeruginosa isolates in this study is not surprising as these genes have been previously reported as the most dominant ESBL and AmpC genes circulating in both the hospital and community settings in Nigeria [2, 3, 31]. The high occurrence of CTX-M-15 ESBL genes in the E. coli, Klebsiella species, and P. aeruginosa isolates has also been reported in China [7]. A higher prevalence of AmpC genes has also been reported in E. coli, Klebsiella species and P. aeruginosa isolates in Switzerland [9]. In Korea, CTX-M-15 was among the most dominant type of ESBL genes recovered from isolates of E. coli [32]. Also in Scandinavia, the CTX-M-15 was the most prevalent ESBL that caused the clonal spread of a multiresistant Klebsiella pneumoniae in that region [33]. Plasmid-mediated ESBL- and AmpC genes are clinically relevant because they have high transmissibility to other bacterial species in the community or hospital environments. The susceptibility and resistance profile of the ESBL- and AmpC-positive E. coli, Klebsiella species, and P. aeruginosa isolate in this study were subjected to antibiogram studies using selected antibiotics. Approximately 78% of the ESBL- and AmpC-positive E. coli, Klebsiella species, and P. aeruginosa isolates investigated in this study were resistant to ceftriaxone, cefotaxime, and doxycycline while 64% were resistant to mupirocin, ceftazidime, and aztreonam. While some of the isolates are notably susceptible to the classes of antibiotics tested in this study, especially the third-generation cephalosporins, carbapenems, and some non-beta-lactam agents, it has been established that E. coli, Klebsiella species, and P. aeruginosa isolates expressing ESBL and AmpC enzymes may develop resistance to some of these antibiotics and thus, should be avoided in the treatment of infections caused by these organisms. Notably, all ESBL- and AmpC-positive E. coli, Klebsiella species, and P. aeruginosa isolates were susceptible to imipenem while approximately 70% of the isolates were susceptible to cefoxitin, sulphamethoxazole-trimethoprim, and colistin sulfate. A critical part of the monitoring, surveillance, and reporting of AMR bacteria in both the community and hospital settings is the establishment of the occurrence rates of ESBL- and AmpC gene-encoding bacteria to inform sustainable infection control practices. Our study has shown that ESBL- and AmpC-producing bacteria are prevalent in the non-hospital environment, particularly in food-producing animals. The evolution and spread of these resistant bacteria can pose a potential risk to the transmission of ESBL- and AmpC-gene-encoding bacteria in the human population in southeast Nigeria. This preliminary study underscores the need to take the monitoring and surveillance of antibiotic-resistant bacteria, particularly those expressing ESBL and AmpC a top priority in southeast Nigeria.
CONCLUSION
AMR has caused a plethora of health crises across the globe, and this is due in part to the intensive (uncontrolled) usage of antibiotics in agricultural practices, animal husbandry, and other clinical and non-clinical applications of antibiotics, especially at sub-inhibitory concentrations. This preliminary study reported the prevalence of AmpC and ESBL genes, particularly the FOX-1 genes and CTX-M-15 genes that mediate the production of AmpC and ESBL enzymes in E. coli, Klebsiella species, and P. aeruginosa that emanated from livestock and poultry milieus in southeast Nigeria. PCR proved to be more precise in detecting ESBL and AmpC in the test bacteria isolates. The presence of AmpC and ESBL-producing bacteria in livestock and poultry milieus is worrying since they can be a key reservoir for the transmission of drug-resistance genes as reported in this study in the human population. These latest findings have led to the identification of key functional genes (including CTX-M-15 and FOX-1) that cause bacterial resistance in southeast Nigeria, and focus attention on the importance of surveillance and monitoring to mitigate the evolution and community transmission of AMR bacteria and ARGs in the environment, as antibiotic therapy could be affected.
Acknowledgments: We thank the staff and management of Enugu State University of Science and Technology (ESUT), Agbani, Nigeria for the administrative support in carrying out this study.
Conflict of interest: None
Financial support: Ethical approval was obtained from the Research and Ethics Committee of the Department of Pharmaceutical Microbiology and Biotechnology, ESUT, Nigeria. All experiment in this study was executed following relevant national and international guidelines.
Ethics statement: None