International Journal of Pharmaceutical and Phytopharmacological Research
ISSN (Print): 2250-1029
ISSN (Online): 2249-6084

In this research, the messenger ribonucleic acid vaccine has proven to be the best vaccine among all others against the Coronavirus disease of 2019, such that the Food and Drug Administration recently gave it full approval. This approval is made possible due to its clinical efficacy, the rare occasion of insertional mutagenesis associated with it, and the absence of anti-vector immunity. Also, its rapid production process gives it an added advantage over conventional DNA vaccines. A literature review was carried out based on available literature online. This review elaborates on the steps involved in the in vitro generation of the messenger ribonucleic acid vaccine. An example is made using a plasmid that contains both the receptor-binding domain found in the spike protein of the Coronavirus and the super folder green fluorescent protein. The use of a viral vector in making these vaccines is also added as a note for additional knowledge.
INTRODUCTION
The production of the messenger ribonucleic acid (mRNA) vaccine via an in vitro process can be done in two ways. It is either done via enzymatic modification, which involves the post-transcriptional modification of the cap and the poly(A) tail of the mRNA or by simply using a template-directed means, also known as co-transcriptional modification [1-4]. The modification via enzymes uses enzymes such as the Guanylyl transferases to catalyze the synthesis of the cap-0 structures during the post-transcriptional modification of mRNA, mainly in Eukaryotes. The successive enzymatic steps required to form cap-0 involves the actions of RNA guanine-N7-methyltransferase and RNA guanylyltransferase in conjunction with that of RNA triphosphatase. The complexity involved in the enzymatic capping limits the regular use of this enzymatic process, as it may not be cost-effective, making the template-directed method a preferred choice [2-4].
This review discusses the in vitro generation of the mRNA vaccine in the context of a plasmid vector as a starting material. It also noted how the viral vector could be used in place of the plasmid vector. For the plasmid vector, the template-directed method of modification has been utilized in the generation of mRNA vaccine, using a plasmid that bears the receptor-binding domain (RBD) of the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) tagged with super folder green fluorescent protein (SFGFP) as an example [5-9]. The summary of the process involved is illustrated with a diagram (Figure 1).
MATERIALS AND METHODS
An online search was carried out on some published works, and relevant literature was used to review the generation of mRNA vaccine via in vitro transcription process and vectors.
RESULTS AND DISCUSSION
Several results as assessed from the literature are presented in Tables 1-4, based on the constituents of the PCR mixture (Table 1), PCR cycling protocol (Table 2), the composition of NTP/cap analog mixture (Table 3), composition of the in vitro transcription (IVT) reaction mixture (Table 4), and functions of the components used in the making of IVT mRNA (Table 5).
Generation of IVT mRNA vaccine: A plasmid vector perspective
Making the IVT mRNA vaccine using a plasmid vector involves producing a quantity of DNA and subsequently making an mRNA out of it. The initial steps of producing the DNA involve the extraction of exogenous DNA templates, the amplification of the linearized DNA, and the purification and testing of this DNA, after which they are transcribed into mRNA. It is worthy to note that after DNA is generated, it is purified using the purification kits and tested for quality control using the DNA gel electrophoresis.
The mRNA produced is also purified using the purification kits and tested for quality control using the RNA gel electrophoresis. They are transfected into the mammalian cells and analyzed using fluorescent microscopy and flow cytometry. These steps are elucidated in the following sections.
Transformation of cells and extraction of exogenous DNA
Plasmids are used in the transformation of cells. In transformation, the plasmids that bear the exogenous DNA are introduced into the E. coli using the super optimal broth with a catabolite repression (SOC) medium. The E. coli is cultured overnight using the LB-Agar. Ampicillin is used to extract the ampicillin-resistant plasmid DNA (pDNA), which bears the gene of the spike protein of the receptor-binding domain (RBD) of the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). It also contains the SFGFP, which served as the selectable marker [5-7]. The plasmids are purified using the plasmid purification kits [5].
Amplification of linearized DNA template (plasmid DNA)
Linearization is the process in which the plasmids are refined. Linearization helps to achieve a definite halt and definite length during the transcription process. This process is essential because of the wild nature of the plasmid DNA (pDNA), which contains impurities. These impurities could usually be traces of the mixture of bacterial genomic DNA and three forms of pDNA (supercoiled, circular, or linear) in variable proportions. The catalytic nature of the RNA polymerases on circular plasmids also necessitates linearization [3, 10-12]. It is important to note that most pDNA template used for in vitro transcription contains at least an open reading frame (ORF), a bacteriophage promoter, a unique blunt-ended restriction site, and optionally a poly(d(A/T)) sequence transcribed into poly(A). This unique blunt-ended restriction site served as the point for the linearization of the plasmid to ensure a definite halt and defined lengths in the transcription process using a restriction enzyme. The most commonly used promoter region is the T7 out of the popular promoter regions T3, T7, or SP6, with their specific phage DNA-dependent RNA polymerase [13-16].
Avci-Adali et al. [5], in their experiment, demonstrated how the DNA template could be linearized and amplified using the constituents of the Polymerase Chain Reaction (PCR) reaction (Table 1). These constituents are passed through the PCR cycling pool (Table 2).
Table 1. Constituents of the PCR mixture
|
Constituent |
Final concentration |
Volume (µL) |
|
Reverse Primer |
0.7 µM |
7 |
|
Forward Primer |
0.7 µM |
7 |
|
5x PCR Buffer |
1x |
20 |
|
5x Q-Solution |
1x |
20 |
|
DNA (Plasmid) |
50 ng / 100µL |
Variable |
|
DNA Polymerase (2.5 U/µl) |
2.5 U |
1 |
|
Nuclease-free water |
|
Variable |
|
Total volume |
|
100 |
The above table shows the quantity of the constituents of the PCR mixture [5].
Table 2. PCR cycling protocol
|
Event |
Cycle number |
Time |
Temp. (°C) |
Aim of event |
|
Initial denaturation step |
1 |
3-7min |
90-100 |
The dsDNA is denatured into single strands to enhance amplification. |
|
3-step cycling: -Denaturation -Annealing -Extension |
2-25 |
40-50 sec 1-3 min 1-3 min |
90-100 50-60 70-75 |
The hydrogen bonds between complementary bases are denatured. The primers are hybridized to the templates. The hybrid is extended via Polymerase. |
|
Final extension step |
26 |
10-15min |
70-75 |
The primer sequences are extended from the 3' end of the primer. |
|
End of cycling |
|
Indefinite |
2-4 |
The cells are preserved. |
The above table shows the events in the PCR cycling protocol and their aims [5, 13-16].
The in vitro transcription (IVT)/ DNase reaction
Avci-Adali et al. [5] in their work, produced the mRNA transcript via a template-directed method using the required constituents (Tables 3 and 4). The NTP/cap analog mixture, which contains a mix of NTP and cap analog, provides the molecular precursors for the RNA. The ATP and GTP are from the MEGAscript T7 Transcription kit, a product of Sigma-Aldrich. The 5-Methylcytidine-5'-Triphosphate (Me-CTP), Pseudouridine-5'-triphosphate (Pseudo-UTP), and the 3´-O-Me-m7G(5')ppp(5')G RNA cap structure analog, also called ARCA, are products of Trilink Biotechnologies. The template DNA and the nucleoside triphosphate concentration determine the yield and the amount of full-length mRNA transcript obtained. The length of CDS and the amount of PCR product used for IVT determine the quantity of the synthesized mRNA. A time-course experiment could be done to decipher the optimal incubation time [5]. During the template-directed co-transcriptional capping of mRNA, the appropriate transcripts are synthesized using 5´-m7G bearing ARCA, which allows their increased translation and correct orientation with the 7-methylated G (m7G) serving as the terminal residue [17-20]. The modified nucleoside triphosphate aims to obviate the in vivo immunogenicity of mRNA vaccines. The need to prevent this immunogenicity could be a reason adjuvant is not added to some mRNA vaccines.
Table 3. Constituents of the NTP/cap analog mixture
|
Constituents |
Stock concentration (mM) |
Final concentration (mM) |
Amount (µL) |
|
ARCA |
10 |
2.5 |
10 |
|
GTP |
75 |
1.875 |
1 |
|
ATP |
75 |
7.5 |
4 |
|
Pseudo-UTP |
100 |
7.5 |
3 |
|
Me-CTP |
100 |
7.5 |
3 |
|
Total volume |
|
|
23 |
The above table shows the NTP/cap analog mixture and its volumes and concentrations [5].
Materials constituting the in vitro transcription (IVT) reaction mixture (Table 4) are RNase Inhibitor, NTP/cap analog mixture, Nuclease-free water, reaction buffer (10x), PCR product, and the T7 RNA polymerase enzyme mix (10x), all serving their respective functions as presented in Table 5. The DNase treatment helps eliminate the genomic DNA, which is now a contaminant in the RNA sample because DNase is an enzyme that selectively degrades the DNA [5, 21, 22]. The PCR products can be purified using the purification kits.
Table 4. Constituents of in vitro transcription (IVT) reaction mixture
|
Constituents |
Final concentration |
Volume (µL) |
|
PCR product |
1 µg |
Variable |
|
NTP/cap analog mixture |
|
23 |
|
Reaction buffer |
1x |
4 |
|
RNase Inhibitor |
40 U |
1 |
|
T7 RNA polymerase enzyme mix |
1x |
4 |
|
Nuclease-free water |
|
Variable |
|
Total volume |
|
40 |
The above table shows the constituents of the IVT reaction mixture with their respective concentration and volume [5].
Purification and quality control of DNA and synthetic mRNA
The DNA purification can be done using the DNA purification kits, while the quality of this purification can be tested using a DNA gel electrophoresis. In the same vein, RNA gel electrophoresis can be used to ascertain the quality of the mRNA transcript produced after the RNA purification kit is used to purify the mRNA. Subsequently, the mRNA transcript is dephosphorylated by treating it with Phosphatases [5, 23].
Transfection and analysis of mRNA containing HEK 293 cells
Transfection is the introduction of exogenous materials into a mammalian cell. The 12-well culture plate is prepared before transfection. Aliquots of Matrigel® (250 µl) and DMEM (200 µl) are brought to -4°C [24-26]. The -4°C is an optimal temperature for the miscibility of the Matrigel and DMEM. After mixing them, the homogenous solution (450 µl) is transferred into DMEM (40 ml), mixed, and distributed into the 12-well plate (1 ml/ well). After which, the plate is kept in the incubator between 2 hours to a maximum of 7 days. Three out of these 12 wells are used to grow the Human embryonic kidney 293 (HEK 293) cells to between 60% and 80% confluency. This growth is done at 37°C, and 5% CO2 within 48 hours. The range of 60% to 80% confluency gives room for other downstream procedures such as flow cytometry and fluorescent microscopy without the cells reaching 100% confluency. This is because at 100% confluency, there would be contact inhibition which makes the cells wither.
The spent DMEM is changed daily until the end of the procedure. The spent DMEM is replaced with Trypsin/EDTA (0.5 ml), and the cells are incubated at 37°C, and 5% CO2 for a maximum of 5 minutes so the cells can be detached. After this time, the Dulbecco's Phosphate-Buffered Saline (1 ml) is used to wash the detached cells. The cells are then incubated in a transfection solution that contains the mRNA. The cationic lipid transfection reagent (0.5 ml) is used at 37°C, and 5% CO2 for 4 hours. Following the transfection of mRNA into the cells, fluorescent microscopy is done. Fluorescent microscopy aims to visualize the cellular structures and examine the expression of proteins and their post-translational modifications [5].
After the fluorescent microscopy, the cells are incubated overnight in DMEM (1 ml). After the incubation, the cells are detached from the 12-well plate using the Trypsin/EDTA (0.5 ml). Subsequently, the cells are washed in DPBS (1 ml) and resuspended in DMEM (10 ml) containing ROCK inhibitor (2µL/10 ml). The resuspended cells are kept in a flow cytometer, and the percentage of the cells expressing SFGFP is analyzed based on their fluorescent intensity, determined by their size, shape, and complexity [5].
Table 5. Critical components needed for the generation of mRNA via the IVT process
|
Name |
Functions |
|
Ampicillin |
Ampicillin inhibits the growth of E. coli, which bears the peptidoglycan cell wall, and facilitates the selective growth of the ampicillin-resistant plasmids [27]. |
|
Transfection solution |
The transfection mixture, such as cationic lipid transfection reagent, helps the mRNA to be delivered into the cytosol. This reagent imparts a polarity similar to the lipid bilayer of the cell on the mRNA, easing the passage of mRNA through the cell (plasma membrane). |
|
SOC medium |
The SOC medium contains glucose, which helps induce a quick adaptation and rapid in vitro growth of the E. coli via the glucose effect phenomenon [28]. |
|
Receptor Binding Domain (RBD) |
RBD is found in the tip of the spike proteins of Coronaviruses. The virus triggers an immune response in humans when it attaches to their ACE 2 receptors via these spikes [4]. |
|
Luria-bertani Agar (LB-Agar) |
The LB-Agar facilitates the growth of pure plasmids of transformed bacterial cells (E. coli). |
|
SFGFP |
SFGFP is a fusion tag for purifying, quantifying, and monitoring proteins in a flow cytometer [5, 7, 29]. |
|
DPBS |
DPBS is physiological saline used to preserve the cells by maintaining an optimal pH. It also helps to annul the hydrolyzing effects of Trypsin/EDTA. |
|
Restriction enzyme |
A restriction enzyme is needed for the linearization of the DNA template to ensure transcripts with defined lengths. It also aids the uni-directional orientation of donor sequence [5, 11, 30]. |
|
RNase inhibitor |
RNase inhibitor annuls ribonucleolytic activity [31-33]. |
|
T7 RNA polymerase |
T7 RNA polymerase is a T7 template-specific polymerase that transcribes the linearized DNA template into mRNA using the nucleoside triphosphates. |
|
Matrigel |
The Matrigel forms the basement membrane needed to support the mammalian (culture) cells. It serves as a surrogate extracellular matrix, which forms the micro-environment for growing and studying the culture cells [24, 25]. |
|
Nuclease-free water |
Nucleases denature the phosphodiester bonds (strands) of nucleic acids. The nuclease-free water is used to supplement the final volume of the reaction mixture to prevent this effect [34, 35]. |
|
DNA Polymerase |
DNA Polymerase speeds up the synthesis of new strands (daughter strands), which are complementary to the strands of the DNA template contained in the reaction mixture found in the PCR [13-15]. |
|
Poly T-tail |
Poly T-tail aids efficient translation in the cells; hence, it is needed as a precursor for mRNA. It is transcribed into a poly-A tail needed to transport mRNA molecules [1, 11, 36, 37]. |
|
Plasmid DNA |
Plasmid DNA is extrachromosomal DNA. It is malleable, self-replicating, and adaptation-efficient. It contains only the coding DNA sequence (exons) needed to be transcribed in bacteria [1-3, 38]. |
|
DMEM |
The DMEM, a nutrient medium, provides nutrients for the mammalian cells. It contains the minimum essential nutrients for the growth of cells. |
|
PCR buffer |
Qiagen posits that PCR buffer contains Factor SB to abrogate the degradation of primers and templates during PCR setup, improving the sensitivity and reliability of high-fidelity PCR. |
|
Q-Solution |
According to Qiagen, Q-Solution regulates the melting characteristics of nucleic acids. It improves the efficiency of the PCR. Efficiency may be attenuated by templates with a high GC content or a high degree of secondary structures. |
|
Reverse primer and Forward primer |
They both flank the target DNA region for amplification. The forward primer (antisense strand) tags the start codon of the template DNA. Reverse primer (sense strand) tags the stop codon on the complementary strand of the DNA [39]. |
|
Poly-A polymerase |
Poly-A polymerase catalyzes the incorporation of poly(A) tail [1]. |
|
Transferases (Guanylyl transferase and 2′-O-methyltransferase) |
Guanylyl transferase and 2′-O-methyltransferase aid enzymatic capping. They confer cap 0 (N7MeGpppN) or cap 1 (N7MeGpppN2′-OMe) structures, respectively. Cap 1 structures modify the mRNA and make it not recognized as foreign by the innate immune system [1-3]. |
|
HEK 293 cells |
The HEK 293 cells are mammalian cells used for in vitro studies. They are generally preferred due to their rapid growth. |
|
DNase |
DNase eliminates the genomic DNA from the RNA samples [21, 22, 40]. |
|
Reaction buffer contains Tris-Hcl and KCl, and sometimes MgCl2 |
Tris-HCl enhances the permeability of the cell membrane. KCl maintains the integrity of the mitochondria, intracellular tonicity, and biosynthesis of nucleic acids. MgCl2 serves as a support for the enzymes, which are biological catalysts [41, 42]. |
|
Anti-Reverse Cap Analog (3´-O-Me-m7G(5')ppp(5')G) |
Anti-Reverse Cap Analog, popularly known as ARCA, is an mRNA cap structure analog incorporated as the first nucleotide of the transcript [1, 2]. The 3´-O-methyl modification ensures mRNA capping is done in the correct orientation and prevents the formation of phosphodiester bonds at this position [1-3]. This leads to enhanced transcription, translation, and stability of mRNA vaccines [17] |
|
Modified nucleosides (Me-CTP and Pseudo-UTP) |
Modified nucleosides such as CTP and Pseudo-UTP substitute the immunogenic native forms of cytidine and uridine, respectively, leading to increased translation, efficient base stacking, improved nuclease stability, and decreased in vivo immunogenicity [1-3, 19]. |
|
ATP and GTP |
ATP and GTP are the molecular precursors of the mRNA. |
|
Trypsin/EDTA solution |
Trypsin optimizes the detachment of the cells from culture plates via hydrolysis. The ethylenediaminetetraacetic acid (EDTA) is an ion-chelator. It keeps the cells in a life-like form during this detachment and enhances hydrolysis [43-47]. |
|
Antarctic phosphatases and inorganic pyrophosphatases |
During the biosynthesis of nucleic acids and proteins, large amounts of phosphates may be produced as by-products, disrupting the completion of these essential physiological reactions. Phosphatases are critical in regulating and reducing the cellular concentration of these phosphates [47-50]. During the generation of IVT mRNA, the phosphatase treatment could be necessary for regulating the excess of phosphorothioate, phosphorothiolate, imidiphosphate, and boranophosphate that may be produced from the use of chemically modified cap analogs [2]. |
|
Rho-kinase (ROCK) Inhibitor |
The ROCK inhibitor fixes the cell. It helps keep the cell viable and maintains its morphology, partly by controlling the stress fibers and focal adhesions. This consequently limits the differentiation and autolysis of cells [51, 52]. |
The above table summarizes the functions of the essential components needed in an in vitro generation of mRNA vaccine and its analysis using cell culture.
|
|
|
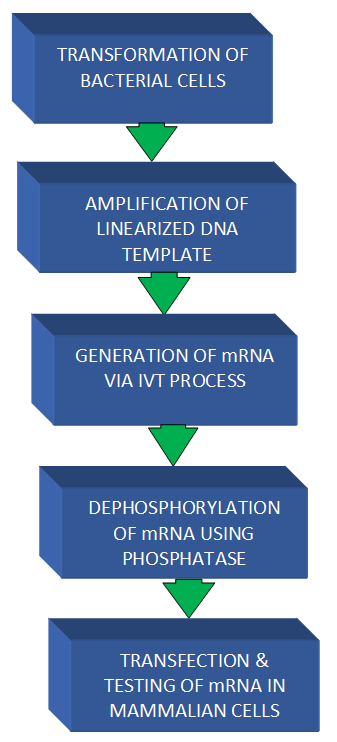
Figure 1. The significant steps in the generation of mRNA vaccine using a plasmid vector. |
The plasmids are extracted from the transformed E. coli and purified using purification kits. The template DNA is linearized, amplified, and purified, after which they are tested via the DNA gel electrophoresis. The IVT mRNA is generated, purified, and its quality tested using the RNA gel electrophoresis. The IVT mRNA is treated with phosphatases, purified, and transfected into the cells. The cells are analyzed using flow cytometry and fluorescent microscopy.
Generation of IVT mRNA vaccine: A viral vector perspective
Although generating an IVT mRNA vaccine using the plasmid vectors as the starting material is the conventional way (Figure 1), there is still an option of using viral vectors in a similar step-wise manner. The difference is that while the conventional way uses a plasmid vector as a starting material, viral vectors are used as the starting material in this instance. Another difference could be seen in their intracytoplasmic effects, as some vaccines are made from viral vectors that can amplify minute doses of the protein. The IVT mRNA made from viral vectors can be broadly classified into non-replicating and self-amplifying vaccines. The former is called non-replicating mRNA (NRM) vaccines because it does not amplify, and the latter, which bears a replicase machinery, is called self-amplifying mRNA (SAM) vaccines because it replicates [1-3, 11]. The modification of the SAM vaccines, also known as the replicon RNA vaccines, gave leeway to the mRNA vaccines called trans-amplifying RNA vaccines [53, 54].
Vaccines made with viral vectors such as the NRM and SAM often use RNA viruses as their vectors. These RNA viral vectors have shown promising results against highly infectious pathogens and many types of cancers. The NRM vaccines can be further classified by their delivery method: direct in vivo injection or ex vivo infusion of dendritic cells into specific human body sites [1]. Both have in common 5′ and 3′ untranslated regions (UTRs), ORF, a cap structure, and a 3′ poly(A) tail. The addition of genetic replication machinery derived from positive sense single-stranded mRNA viruses predominantly from Flavivirus such as the Kunjin virus, and the Alphaviruses such as Sindbis virus and Semliki Forest virus, distinguished SAM vaccines. Generally, the ORF that encodes the structural proteins of the virus are substituted by the gene of interest (GoI), and the RNA-dependent RNA polymerase of the virus is used to facilitate the amplification of the replicon construct in the cytosol [1, 11].
The presence of the replicase machinery in the SAM vaccines obviates the need for an adjuvant. This is because the replicase machinery amplifies the dose of these vaccines once in the cytoplasm. Hence a low dose could be amplified to a large dose intracellularly, making it sufficient to confer the needed long-lasting immunity. This is why the SAM vaccines are more efficient as prophylaxis for infectious diseases. The NRM vaccines, which are mainly used as an anti-cancer vaccine, do not possess replicase machinery. Thus, it requires adjuvants such as the key-hole limpet hemocyanin [1, 40, 55].
The hallmark of the SAM mRNA vaccine is seen in its use as prophylaxis against infectious diseases. A use whose clinical efficacy is determined by the efficiency and structure of the 5′ cap; the UTR length, structure, and regulatory elements; the modification of the coding sequence; the poly-A-tail properties; and the purity [1, 3]. The SAM vaccines used against Coronavirus disease of 2019 (COVID-19) are not associated with anti-vector immunity, a type of immunity against the viral vectors made of Adenovirus, seen in those previously exposed to this virus [56, 57]. This anti-vector immunity could render these vaccines less potent. Insertional mutagenesis is a phenomenon by which sequences of exogenous nucleic acids integrate within the genome of the host cell, leading to deregulation of genes adjacent to the insertion site and, in some cases, disruption of cellular phenotype [58]. The mRNA vaccine is rarely associated with insertional mutagenesis. Also, the mRNA vaccine is non-infectious due to the rigorous purifying and refining processes involved in its production. These purification processes also circumvent the need for antimicrobials such as Ampicillin.
Despite the massive gains made with the mRNA vaccine, its major limitation is its inability to contend with viruses with high mutating capacity because it is mainly composed of modified nucleosides, a type of synthetic nucleotide. Mutation comes with changes in the nucleotide sequence, which influences the strains. Subtle genetic changes in some viruses may elicit mutations that could lower the potency of the vaccine, while drastic genetic changes may occur via recombination and could make the vaccines impotent [59-63]. The tendency for Coronaviruses to display trans-species movement and adaptation could invariably influence the efficacy of the mRNA vaccine in line with the species of the host, season of the year, and geographical location [60-64]. A typical scenario is seen in the ongoing pandemic where the efficacy of the vaccine drops as new strains of SARS-CoV-2 emerges. According to Pfizer, the South African variant of SARS-CoV-2 (B.1.351) overwhelmed the efficacy of the mRNA vaccine seen in South Africa by about 70%, as communicated by Consumer News and Business Channel [64].
CONCLUSION
Producing a potent vaccine involves a rigorous process in which cell culture or laboratory animals are utilized in analyzing the quality of the end-product. This rigorous process requires sound knowledge and experience in clinical immunology and its associated molecular techniques. Hence, the governments of various countries and their respective public health agencies could engage those highly experienced in this field to ramp up the production of these vaccines. This would go a long way to bring the pandemic to an end, despite the varying efficacy of the vaccines among people, a variation partly attributed to pharmacogenomics.
Acknowledgments: Special thanks to all the resilient, law-abiding, and cooperative Nigerians who contributed to making Nigeria the fourth safest country during the COVID-19 pandemic.
Conflict of interest: None
Financial support: None
Ethics statement: None