International Journal of Pharmaceutical and Phytopharmacological Research
ISSN (Print): 2250-1029
ISSN (Online): 2249-6084
|
Evaluation of Immune Response Following Experimental Diphtheria-tetanus Vaccine Administration in Mice Model
Azadeh Fardipoor1, Taghi Zahraei Salehi2*, Majid Tebyanian3, Ramak Yahyaraeyat2 |
|
1 Department of Pathobiology, Science and Research Branch, Islamic Azad University, Tehran, Iran; 2 Department of Microbiology and Immunology, Faculty of Veterinary Medicine, University of Tehran, Tehran, Iran; 3 Department of Biotechnology, Razi Vaccine and Serum Research Institute, Agricultural Research Education and Extension Organization (AREEO), Karaj, Iran. |
ABSTRACT
According to the lack of a comprehensive non-clinical study for evaluating the national diphtheria-tetanus (DT) vaccine, we developed an experimental DT vaccine for comparing immunogenicity and efficacy of the DT vaccine with the international standard vaccine. Experimental vaccines were formulated as an adjuvant and non-adjuvant models using diphtheria and tetanus antigens with aluminum sulfate (alum) adjuvant. We used a unique formulation of the vaccine that was associated with greater antigen uptake on the alum gel. The quality control tests including identity (flocculation), adsorption, and potency tests have been performed to evaluate the potency of the vaccines. The pattern of the humoral immune response was calculated and plotted by an indirect ELISA method following immunization in mice model. The immunogenicity of the diphtheria-tetanus vaccine was investigated with serum neutralization (SN) and ELISA tests. Our findings indicated that the experimental DT vaccine was capable of inducing a long-term humoral immune response in mice and could induce a strong humoral response after the first injection. These results demonstrated the efficacy of the above vaccine in monovalent and divalent forms. A significant difference was observed between vaccines formulated with and without adjuvant. These findings indicated adjuvants can stimulate stronger humoral immunity against the target antigen and can be effective in prolonging immunity. To conclude, the experimental DT vaccine has efficacy and is capable to stimulate humoral immune responses.
|
Key Words: Diphtheria-Tetanus vaccine, Immunogenicity, ELISA, Immune response. Vaccination during the first year of life elevates the level of antibodies, which usually remain sufficient until puberty, therefore adults should receive a booster dose of the vaccine [1]. Vaccination against bacterial infection is a low-cost method that could be used for preventing fatal diseases. According to the worldwide vaccination program, the prevalence of many hazardous diseases among infants, children, and adults has been extremely reduced [2, 3]. DT vaccine is an alum adjuvant-based toxoid killed vaccine containing diphtheria and tetanus inactivated toxin that is used for immunization of children against these dangerous infections. Diphtheria is a local mucosal infection caused by the Corynebacterium diphtheriae. This bacterium produces a toxic protein that causes myocarditis, swelling of nervous tissue and has destructive effects on other organs [4]. Tetanus is an acute and fatal disease caused by Clostridium tetani. After the colonization of Clostridium tetani in cutaneous wounds, it secretes a lethal toxin, called tetanus spasming, which can be reached to the nervous system and leading to clinical manifestations of tetanus disease [5]. Regardless of the global vaccination against these fatal diseases, the World Health Organization (WHO) reports indicate many new cases and mortality of this infection. Therefore, control of diphtheria and tetanus has been targeted in healthy programs [6]. DT vaccine is used in Iran to immunize infants and children aged between 2 months and 6 years who are sensitive to the diphtheria-tetanus-pertussis (DTP) vaccine. In general, the vaccination strategy varies from country to country, and each country must continuously review the epidemiology of diseases and adjust its national vaccination policy and program [7]. Tetanus disease is a rare disease in developed countries due to effective vaccination programs, but it is still reported in adults and populations with inadequate and effective vaccination [8, 9]. Infant vaccination programs for the DT vaccine began in 1940 [10]. Vaccination against diphtheria and tetanus has been applied in Iran since 1950 [11]. Effective vaccines should be able to create a reasonable protective immune response. Therefore, the potency and efficacy of vaccines must be evaluated by appropriate immunological tests [12]. According to the previous reports, in the case of the DT vaccine, as a toxoid killed vaccine, the evaluation of humoral immunity is important [12]. Another important factor that must be held in the production of effective vaccines, is to produce an immune memory. Primary vaccination elicits both protective immunity and immunological memory. Immunological memory provides a rapid and strong immune response upon re-exposure to the pathogen [13]. Vaccine immunogenicity would be evaluated in proper animal models. Paramount data will be achieved by immunogenicity studies in the animals concerning adjuvant formulations optimization and antigen evaluation such as the ability to induce functional antibodies [14]. Preclinical investigations can evaluate the combination of adjuvant and antigen as formulated for clinical use [14]. In the present study, according to the lack of a comprehensive non-clinical study for evaluating the national DT vaccine, we developed an experimental DT vaccine for comparing immunogenicity and efficacy of the national DT vaccine with the international standard vaccine. MATERIALS AND METHODS Vaccine formulation The experimental vaccines using diphtheria and tetanus antigens (monovalent and bivalent) were formulated with and without aluminum phosphate (alum) adjuvant [15]. Each experimental vaccine (0.5 ml) contained 20 Lf (Limit of Flocculation) of diphtheria antigen, 6 Lf from tetanus antigen, and 20 mg aluminum. Each dose (0.5 mL) of the National Institute for Biological Standards and Control (NIBSC) diphtheria vaccine standard (22/27) and tetanus vaccine standard (98/552) containing 20 Lf of diphtheria toxoid and 6 Lf of tetanus toxoid, respectively.
Vaccine quality control After formulating the experimental vaccines, quality control tests including identity test (flocculation test) to characterizing antigen content of the vaccines, adsorption test for determining the percentage of adsorption of antigens in aluminum phosphate gel and potency test to evaluate the potency of the vaccines were performed.
Potency test The vaccine's ability against related infection was evaluated by the potency test. Briefly, ten Pirbright guinea pigs weighing 250 to 350 g were immunized with 0.75 ml (1/2 total human dose) vaccine subcutaneously. After 6 weeks, blood was collected and serum was separated and then incubated for 30 min. at 56 ° C for complement inactivation and stored at -20 ° C until tested [15, 16]. The animals were cared for based on the recommendation of the ethical committee of Islamic Azad University, Science and Research Branch following the ‘Guide for Care and Use of Laboratory Animals’ National Institutes of Health (NIH) US publication 86-23, revised 1985 and the protocols were approved by this committee.
Serum neutralization (SN) test This assay was performed to find out the antibody levels and function in the presence of dependent antigens. A range of dilutions (from 0.1 up to 1) of serum was added to ten tubes and the final volume was set with peptone water on 4 ml for tetanus and 1 ml for diphtheria. Subsequently 1 ml of tetanus toxin, (5 L + / 10) (L+/10 refers to the smallest amount of toxin causing paralysis when mixed with 0.1 IU of reference antitoxin) [15]. and diphtheria toxin (10 Lr /1000) (Lr/1000 test dose refers to the smallest amount of toxin causing an erythematous lesion despite the presence of 0.001 IU reference antitoxin) [15]. were added to related tubes. After 1 h incubation at 25° C the content of each tube with tetanus antigen was subcutaneously injected into 3 NMRI mice (0.5 ml) and the animal walking pattern was recorded for 5 days. Then, 0.2 ml of each diphtheria related tubes were injected intradermally into the two Dutch rabbits and they were monitored up to 3 days for any symptoms such as necrosis, ulcer, and redness [15].
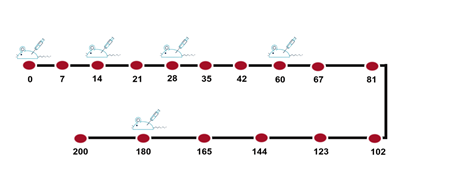
Vaccine identity test (Flocculation test) This assay, known as Ramon assay, is based on the observation of a macroscopic flocculation complex. The levels of diphtheria and tetanus antigens in formulated vaccines were determined with this test. For this matter, sodium citrate was added to the vaccine with a final concentration of 7% and incubated at 37 ° C for 20 hours. Then, the standard working of diphtheria and tetanus antitoxin (100 Lf / ml) was added at 0.03 ml intervals to 10 tubes. Then 1 ml of the vaccine that opened its alum gel was added to each tube and they were transferred at 45° C in a water bath. After that, the contents of each tube were examined for any flocculation turbidity in front of the light bulb until the first reaction was observed. The 1st tube in which flocculation appears is used to determine the Lf value of the sample. Then, we calculated the vaccine titer by antigen concentration (Lf/ml): volume of antitoxin in the first tube that flocculation was observed (ml) × 100 [15]. Vaccine adsorption test Adsorption test was carried out for measuring antigens adsorption to aluminum phosphate gel. We put the vaccine in non-moving conditions to precipitate the gel and then centrifuged the vaccine supernatant (2000 rpm/10 min). Ten tubes were labeled and different amounts of standard diphtheria and tetanus antitoxin (100 lf / ml) at 0.03 ml intervals were added to each one and the final volume set on 1 ml with PBS. Then, 4 ml of vaccine supernatant was added and incubated at 45 ° C in a water bath. The contents of each tube were examined for any flocculation reaction in front of the light bulb until the first flocculation was observed [15]. Immunization protocol Diphtheria antigen and tetanus antigen were injected into mice in 4 stages for the preparation of hyperimmune serum. For immunization with experimental vaccines, a total of 90 female Balb/c mice, 6 to 8 weeks of age, were randomly divided into 9 groups including 10 mice in each group [17]. According to figure 1, 100 µl of vaccines were injected by the intramuscular route 5 times (days 0, 14, 28, 60, and 180) to the mentioned groups [18, 19]. Serum was separated after every 14 steps and kept at -20 ° C until ELISA test [15, 20].
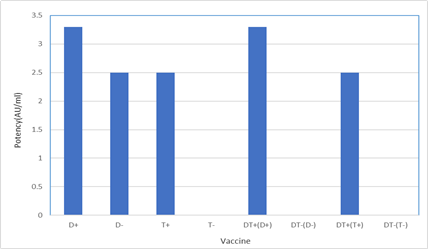
Figure 1: The immunization schedule 100 µl of vaccines were injected by intramuscular route in 5 steps (days 0, 14, 28, 60, and 180) to 9 groups of female Balb/c mice, 6 to 8 weeks of age, and serum was separated after 14 steps. The circles represented the bleeding times. Evaluation of humoral immune response against diphtheria and tetanus antigens using ELISA The levels of specific antibodies against diphtheria and tetanus antigens were evaluated by indirect ELISA method. To do this, the ELISA set-up method in the laboratory was used. The optimum concentration of antigen and antibodies were determined by checkerboard analysis. After obtaining the best concentration of serum and antigen and conjugated antibody using the checkerboard, the ELISA test was performed to check the antibody titers after each blood sampling step. The 96-well plates were coated with 100 μl of diphtheria (ng/well) and tetanus (ng/well) antigens diluted in carbonate-bicarbonate buffer pH 9.6, after overnight incubation at 4 °C. The wells were blocked with 300 μl of 3% bovine serum albumin (BSA) in PBS containing 0.05 % Tween-20 (PBS-T) and incubated for 1 h at 37 °C. The wells were washed 3 times with PBS-T (washing buffer) and the serum samples (1:100 diluted) were added to the wells in question. Then plates were incubated for 1 h at 37 °C and, they were washed for 4 times and 100 μl of HRP-conjugated rabbit anti-mouse IgG (1:25000 diluted) were added to each well and followed by 1 h incubation at 37 °C. Following 5 times of washing steps, 100 μl of Tetramethylbenzidine (TMB) substrate was added to each well and incubated for 15 minutes at room temperature and darkness. Finally, the reaction was stopped by a sulfuric acid solution and the optical absorbance optical density (OD) of the samples was measured at 450 nm with ELISA reader (Bio-Tek). For each sample, two ELISA wells were assayed in each study group to evaluate the antibody level in a binary manner. The antibody curve was obtained from vaccine injection into mice (humoral immune response kinetics) was calculated and plotted by ELISA. RESULTS The potency of vaccines (serum neutralization) Vaccine ability against infectious agents has been evaluated by the potency test (Table 1 and figure 2). A comparison of the SN test results demonstrates that monovalent and divalent diphtheria and tetanus vaccines with adjuvant (D+, T+, DT+(D+), DT+(T+)) produced high-affinity antibodies which were able to neutralize diphtheria and tetanus toxins. About all of them, neutralizing antibody levels were according to the internationally accepted range (≥ 2 AU/ml). Also, monovalent tetanus vaccine and divalent diphtheria and tetanus vaccines without adjuvant (T-, DT-(D-), DT-(T-)) produced low-affinity antibodies which were unable to neutralize diphtheria and tetanus toxins that their neutralizing antibody levels were zero.
Table 1: Comparison of experimental vaccine serum neutralization (SN) test
Groups of ten guinea pigs were immunized with the experimental vaccines monovalent and bivalent were formulated with and without adjuvant. Diphtheria and tetanus antitoxin concentrations of guinea pig serum determined by serum neutralization (SN) test to find out the antibody levels to diphtheria and tetanus antigens.
Figure 2: Potency of vaccines (AU/ml) (+) with adjuvant; (-) without adjuvant
Identity and adsorption of vaccines After the formulation of a vaccine with alum adjuvant, the identity and adsorption tests were performed. The results demonstrated that vaccine titer and absorption rates were 40 Lf/ml and 90% for diphtheria, 12 Lf/ml, and 80% for tetanus respectively.
Checkerboard ELISA Based on the results of the checkerboard ELISA, the most appropriate concentration for diphtheria antigen is 25 ng / well, for tetanus antigen is 15 ng / well, the most appropriate dilution for diphtheria and tetanus serum is 1/100 and for conjugate is 1/5000.
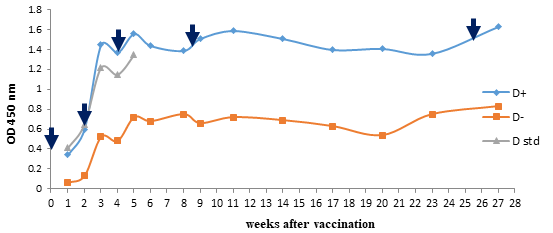
Kinetics of antibody response to vaccines after immunization Monovalent diphtheria vaccine The antibody responses to the monovalent diphtheria vaccine have been demonstrated in Figure 3. There was no significant difference in immunogenicity of experimental vaccine and standard diphtheria vaccine (p<0.001), whereas a significant difference was observed in the monovalent diphtheria vaccine which has been formulated with and without adjuvant. After immunization of mice with a monovalent diphtheria vaccine with adjuvant, the immune response was boosted significantly following the first reminder that repeated after second and third booster injection and reached its highest level in the fifth week of immunization.
Figure 3: Comparison of experimental monovalent diphtheria vaccine with and without adjuvant and standard diphtheria vaccine. Diphtheria antitoxin levels in mice at various time intervals after vaccination with experimental and standard monovalent diphtheria vaccine determined with indirect ELISA assay. A significant difference was seen in the monovalent diphtheria vaccine with and without adjuvant. The injections administered at 0, 14, 28, 60 and180 days which display in the graph with arrows (↓). (+) with adjuvant; (-) without adjuvant
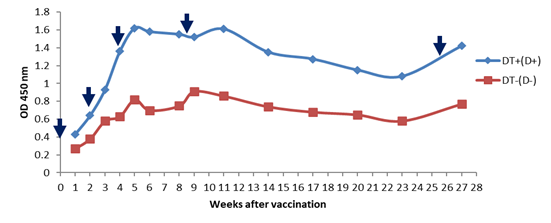
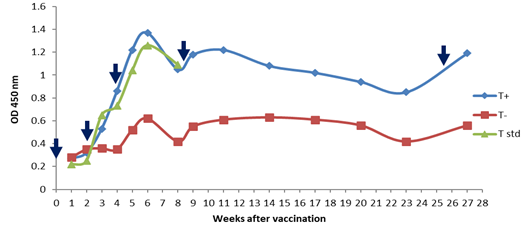
DT vaccine (Diphtheria antigen) The antibody responses to diphtheria antigen after immunization with DT vaccine were displayed in Figure 4. There was a significant difference between the groups which received the vaccine with and without the adjuvant. In the group receiving diphtheria vaccine with adjuvant, the immune response, such as monovalent diphtheria with adjuvant, was elevated sharply after the first booster injection. After the second reminder injection, a mild elevation was observed due to the highest level of the immune response. In the DT vaccine without adjuvant, the immune response was raised slightly after the first and second reminders, but no increase in the immune response was detected after the third booster.
Figure 4: Comparison of experimental diphtheria-tetanus vaccine with and without adjuvant (diphtheria antigen).
Diphtheria antitoxin levels in mice at various time intervals after vaccination with the experimental bivalent diphtheria-tetanus vaccine with and without adjuvant determined with indirect ELISA assay. A significant difference was seen in the diphtheria-tetanus vaccine (DT) with and without adjuvant. The injections administered at 0, 14, 28, 60 and180 days which display in the graph with arrows (↓). (+) with adjuvant; (-) without adjuvant Monovalent tetanus vaccine As shown in Figure 5, there was no significant difference in the immunogenicity of the experimental vaccine and standard tetanus vaccine. However, a significant difference was observed between adjuvant and non-adjuvant monovalent tetanus vaccines after the first reminder injection, the immune response in the adjuvant vaccine model was boosted significantly, whereas in the non-adjuvant model there was no elevation in the immune response. The highest antibody responses were detected during the sixth week. After the sixth week, the immune response to the adjuvant vaccine was declined sharply. Tetanus antitoxin levels in mice at various time intervals after vaccination with experimental and standard monovalent tetanus vaccine determined with indirect ELISA assay. A significant difference was seen in the monovalent tetanus vaccine with and without adjuvant. The injections administered at 0, 14, 28, 60 and180 days which display in the graph with arrows (↓). (+) with adjuvant; (-) without adjuvant DT vaccine (Tetanus antigen) Regarding Figure 6, the DT vaccine with adjuvant has greater immune responses which have been sharply raised after the first booster injection. In the group receiving the DT vaccine without adjuvant, the immune response was raised steeply after the second reminder. However, a weaker elevation in the immune response was observed after the third and fourth reminders injection. Tetanus antitoxin levels in mice at various time intervals after vaccination with the experimental bivalent diphtheria-tetanus vaccine with and without adjuvant determined with indirect ELISA assay. A significant difference was seen in the diphtheria-tetanus vaccine (DT) with and without adjuvant. The injections administered at 0, 14, 28, 60 and180 days which display in the graph with arrows (↓). (+) with adjuvant; (-) without adjuvant. DISCUSSION
Vaccination against three diphtheria, tetanus, and pertussis diseases has been established by WHO since the 1940s as part of its expanded program immunization (EPI) and most countries follow this plan [21]. The result of this program is to reduce children and infant mortality from three fatal diphtheria, tetanus, and pertussis diseases [22]. Up to our knowledge, there is not any comprehensive study in Iran that covers all aspects of immune responses following the administration of DT vaccine containing diphtheria and tetanus antigen. In the present study, experimental diphtheria and tetanus vaccine was formulated and the pattern of immune responses following immunization was examined in mice model. According to our findings, the formulated vaccine could induce a strong humoral response after the first injection. Our data also demonstrated the strongest immune response was detected in the fifth week regarding the monovalent diphtheria vaccine with adjuvant. Indeed, most immune stimulation occurred after the first booster injection, and subsequent reminders only stimulate the immune system to maintain the immune response for up to 28 weeks. The results of a similar study indicated a significant elevation in antibody titers after the first booster dose [23]. Interestingly, Wari et al reported that the highest antibody titer against diphtheria was observed in the fifth week after immunization, which is in agreement with the result of the present study [24]. Likewise, the anti-diphtheria antibody has been raised after the first booster injection of DT vaccine with adjuvant and reached its highest level with a slight slope. This is similar to other studies that represented the DT vaccine provides very good immunity against diphtheria antigen. In other investigations, a significant elevation in antibody titers after the first dose of diphtheria vaccine reminder injection was observed [25]. The findings of the study by Larsenk et al demonstrated that the DT vaccine provides very good immunity against diphtheria and is consistent with our results [26]. In the case of the monovalent tetanus vaccine, the levels of specific antibodies have been increased after the first reminder and reached its highest level on week 6 with a mild grade. However, the immune response was declined by adjuvant and non-adjuvant vaccine after the sixth week suggesting that re-administration of the reminder dose is necessary. Similarly, the other studies advised for tetanus booster doses for maintaining immune responses [27]. Our findings indicate that our experimental DT vaccine is capable of inducing a long-term humoral immune response in mice and this is in agreement with similar reports that emphasized administration of DT vaccines with appropriate reminder doses, which could induce a long-term humoral immune response [28]. Initial immunization creates protective immunity, and immune memory induces a strong and specific immune response by injecting a booster dose and providing long-term protection even in people with low levels of antibody [13]. On the other hand, the short interval between reminder doses may lead to unnecessary vaccination of immunocompromised individuals and does not lead to long-lasting immunity [13, 29]. Bitragunta et al investigated the immunogenicity of a dT vaccine in primary school children in Hyderabad India. They found that a dose of dT vaccine as a booster in primary school children produced more than 96% protective antibody. It has been reported that multiple doses of tetanus vaccine reminders may increase serum tetanus antibody titers but may not lead to an effective immune response [30]. In a parallel study, Silva et al evaluated the short-term and long-term immune response in mice after immunization with the diphtheria vaccine، and concluded that the DTP vaccine produced a long-lasting immune response against diphtheria [18]. In the present study here, in all experimental vaccines, a significant difference between formulated vaccines with and without adjuvant was observed. These findings indicate adjuvants can stimulate stronger humoral immunity against the target antigen. In another study, Alshanqiti et al reported that there was a large difference in immunogenicity between the tetanus vaccine formulated with alum adjuvant and non-adjuvant vaccine [31]. Using adjuvants can be effective in prolonging immunity (creating long-lasting immunity), activating greater numbers of immune memory cells, and raising the amount and affinity of antibody production [32]. Antibody elevation in adjuvant groups indicates activation of active memory cells following vaccination and this is not detected in the non-adjuvant vaccine [33]. A comparison of the SN test results between the DT vaccine with and without adjuvant indicates that in the non-adjuvant groups, the antibodies had very low affinity and could not neutralize diphtheria and tetanus toxin, despite the acceptable antibody titers confirmed in the ELISA test. This confirms the role of adjuvant in enhancing antibody affinity [34]. In a study, the immunogenicity of diphtheria vaccine was investigated with SN and ELISA tests and indicated that serum neutralization test could especially detect neutralizing or protecting antibodies but both the neutralizing and non-neutralizing antibodies were detected in the ELISA method because ELISA is a specific binding assay which does not distinguish between neutralizing antibodies from non-neutralizing antibodies [35]. This is an important point that has been highlighted in our study. CONCLUSION To sum up, the present study demonstrated that the experimental DT vaccine is capable to stimulate humoral immune responses. The results indicated that immune responses shift toward Th2 and humoral immune responses. The kinetic antibody responses following the first injection represented an increasing gradient after each booster dose and immune response has been stable for more than 100 days. The protective potential of antibodies was confirmed in the SN test. These results demonstrate the efficacy of the above vaccine in monovalent and divalent forms and endorse the ability of alum adjuvant for enhancing humoral immune responses.
Funding This research received no external funding. ACKNOWLEDGMENTS The authors would like to thank all of the personnel and colleagues in the Razi Vaccine and Serum Research Institute, especially departments of Human Vaccine and Serum, Immunology, and quality control. Conflicts of interest The authors have no conflict of interest. REFERENCES
|
||||||||||||||||||||||||||||||||||||